INTRODUCTION
Sleep accounts for approximately one-third of human life and has been an essential part of human survival [1]. However, sleep has been regarded as a passive state or physiological phenomenon, and the significance of sleep has not been recognized. In particular, appropriate sleep duration is significant to maintaining vigilance during waking. Sleep deficiency is clinically associated with a variety of sleep disorders, such as insomnia and insufficient sleep syndrome [2]. Also, short sleep duration can increase the risk of various consequences, including cardiovascular disorders, metabolic diseases, and mortality [3-5].
An allergy is characterized by overactive responses of the immune system resulting from a specific allergen [6]. Allergic disease is clinically related to several chronic inflammatory disorders such as allergic rhinitis, atopic dermatitis, and asthma [7].
A few studies have investigated the link between sleep deprivation and allergic inflammatory response in animal models [8,9]. However, it is not fully known about the relationship between sleep deprivation and allergic inflammatory response. We hypothesize that insufficient sleep could cause the deterioration of the allergic reaction. Thus, the purpose of this study was to determine the possibility that sleep deprivation is associated with inflammatory response in rats.
METHODS
Animals
Male Wistar rats of age 14 weeks were used for the experiment and randomly assigned to a sleep deprivation [sleep deprivation (SD) group, n = 3] or a control group [healthy sleep (HS) group, n = 3). They were in a controlled environment, with a 12-hour light-dark cycle and free access to food and water. All rats used in the study were handled according to the protocol approved by the Institutional Animal Care and Use Committee of Inha University Hospital (INHA 181207-605).
Animals & Protocol for Sleep Deprivation
A modified multiple platform model was used to induce sleep deprivation (Fig. 1) [10]. To be brief, we installed several 6.5 cm diameter cylinders inside the cage and filled the water up to 1 cm below the height of the cylinder. The rats could move freely on the platform and to feed and intake water. However, when rats sleep, they lose their center on the platform and fall into the water so that we could induce rat sleep deprivation. We also set up a ‘platform cage control group.’ In this group, we used the same cage and filled the water equally up to 1 cm below the cylinder. However, because the platform was 14 cm in diameter (much broader compared to the SD group), the rats could sleep on the platform. We exposed each rat continuously for 72 hours under SD or HS conditions and sacrificed them immediately after exposure. Before and after 72 hours of HS or SD exposure, we measured the weight of each rat and calculated the change.
Measurement of Serum Levels of Interleukin-1Beta and Corticosterone
Immediately after the exposure to SD or HS, the rats were sacrificed. We collected blood by the aortic puncture technique from the abdominal aorta. We then centrifuged the whole blood for 30 minutes at 4°C, 13,000 × g. The supernatant was immediately stored at -80°C until future analysis. After diluting the serum to 1:50, we measured the serum level of interleukin-1beta (IL-1β, R&D Systems, MN, USA) and corticosterone (Abcam, Cambridge, UK) using enzyme-linked immunosorbent assay, according to the protocol provided by the manufacturer.
Collection of Broncho-Alveolar Lavage Fluid
To collect broncho-alveolar lavage fluid (BALF), we cannulated the trachea using the polyethylene tube. We lavaged the lung with Hank’s balanced salt solution (Thermo Fisher Scientific, Waltham, MA, USA). We centrifuged the lavage fluid at 4°C for 15 minutes, at 3,000 × g. The pellet was immediately suspended in physiologic saline for cell counting. We determined the total cell numbers in duplicate using a hemocytometer. We centrifuged an aliquot of 100–200 μL in a Cytospin 2 cytocentrifuge (Shandon Scientific, Pittsburgh, PA, USA). We determined the differential counts of eosinophils, neutrophils, and lymphocytes from the centrifuged preparation, which were stained using the Diff-Quik stain kit (Sysmex Corp., Kobe, Japan) by counting > 500 cells from each sample at × 200 of magnification. The representative photograph of BALF neutrophils are shown in Fig. 2.
Histopathologic Examination
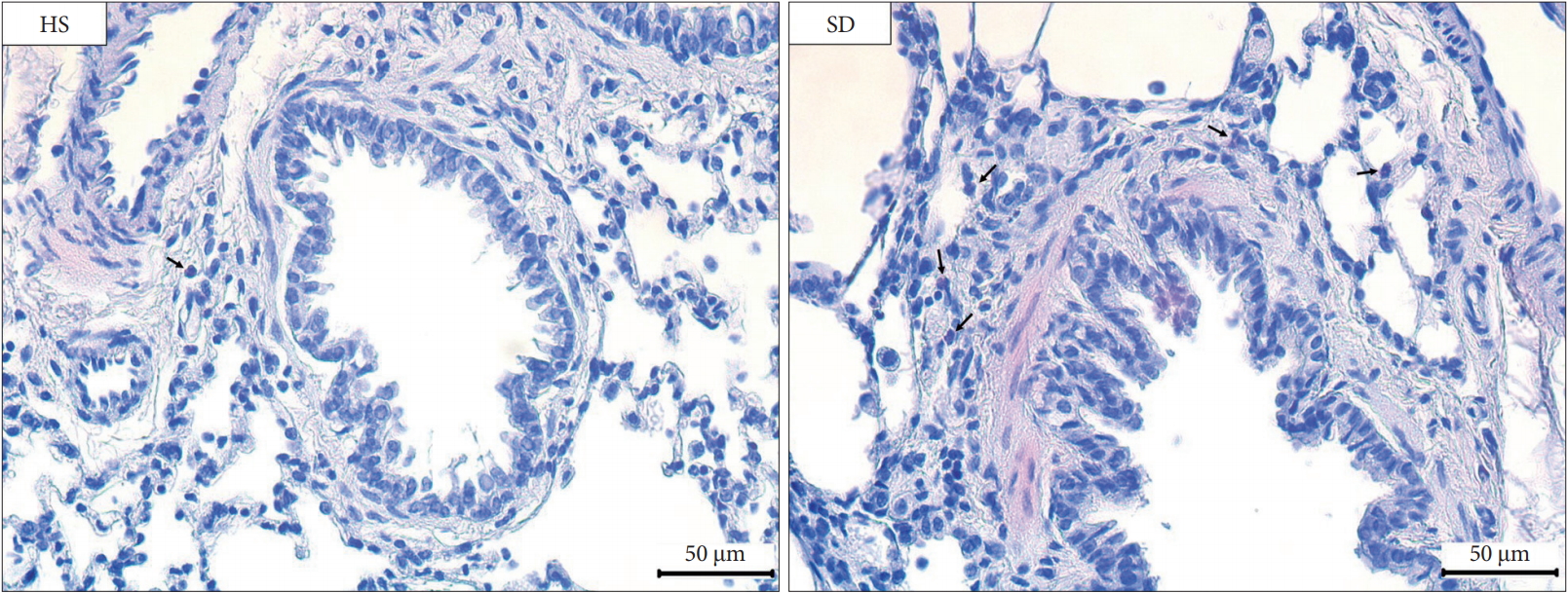
We fixed tissue specimens of the lung in 4% paraformaldehyde for 24 hours. We washed the lung tissues using deionized water, and then, embedded them in paraffin. We stained the tissue sections (of 3 μm thickness) using hematoxylin & eosin. We examined the total counts of eosinophils in pulmonary parenchyma (peri-bronchiolar infiltration).
Statistical Analysis
We used the SPSS 22.0 program (IBM Corp., Armonk, NY, USA) for the statistical analysis. We adopted the nonparametric tests, such as the Kruskal-Wallis test to compare the change of body weight, serum IL-1, corticosterone, the number of inflammatory cells in BALF and the counts of eosinophils in lung parenchyma. A p-value < 0.05 was statistically significant.
RESULTS
After 72 hours, rats in the HS group had some gain of body weight (4.3 ± 2.0 g). Conversely, rats belonging to the SD group showed significant decrease in weight after sleep deprivation (-20.2 ± 2.5 g, p = 0.005).
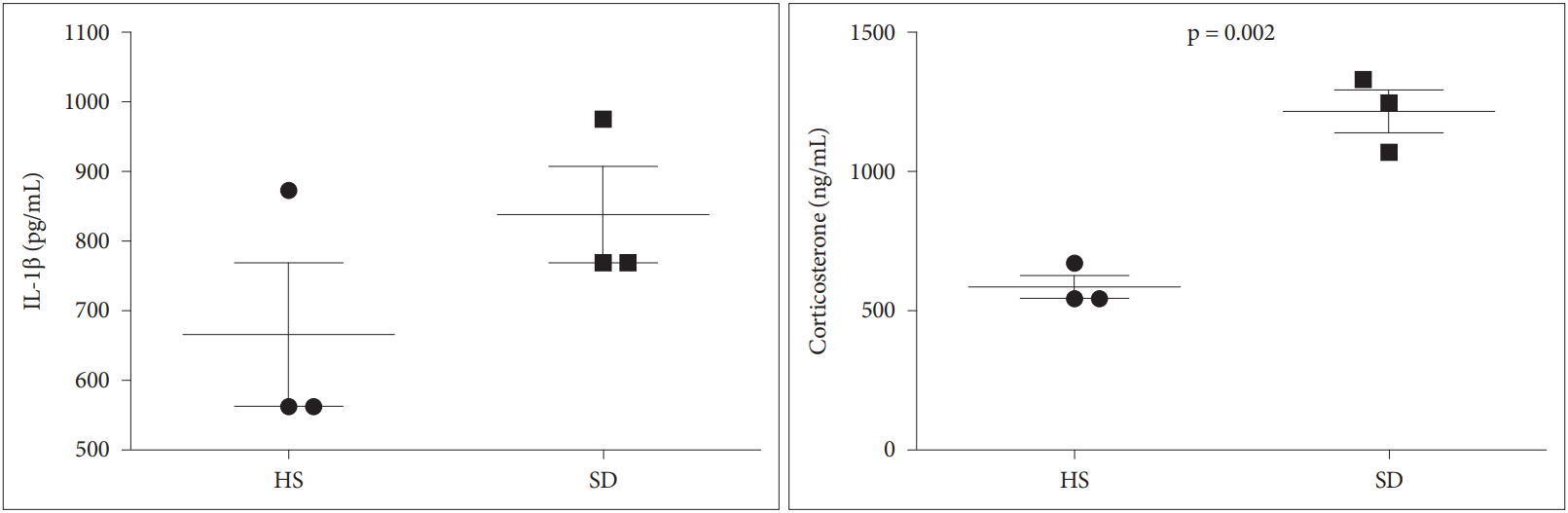
Compared with the HS group, the SD group showed increase in serum IL-1β and corticosterone, and the latter showed a statistically significant difference (p = 0.002 for corticosterone) (Fig. 3).
When comparing the number of inflammatory cells in BALF of the HS group and the SD group, the SD group showed more neutrophil and eosinophil count. However, this did not have statistical significance (p > 0.05 for eosinophil, neutrophil and lymphocyte) (Fig. 4).
Comparing the difference of the pulmonary parenchyma according to the sleep patterns between the groups, the SD group showed increased peri-bronchiolar infiltration of eosinophils (Fig. 5). Comparing the count of eosinophils in 1 mm2 of lung parenchyma, we could note that the SD group had significantly increased infiltration of eosinophils compared to the HS group (the HS group: 4.5 ± 0.9 × 103/mm2 versus the SD group: 16.7 ± 5.2 × 103/mm2, p = 0.017).
DISCUSSION
Various allergy-related parameters including IL-1β, corticosterone, the number of inflammatory cells in BALF, and the histopathology of the pulmonary parenchyma were evaluated in this study to determine the relationship between sleep deprivation and allergic inflammation through animal experiments. The outcomes of this study provided that the SD group had increase in serum IL-1β, corticosterone, the number of inflammatory cells (e.g., neutrophil and eosinophil) in BALF, and peribronchiolar infiltration of inflammatory cells compared to the HS group. Diverse mechanisms may be presumed for sleep deprivation to affect the exacerbation of allergic reactions. As is well known, sleep deprivation is associated with chronic inflammation, nervous or hormonal imbalance, and immunologic change [8,9,11,12]. These can cause or worsen allergic responses. Irie et al. [8] evaluated the influence of REM sleep deprivation using the classical platform model on immediate and late asthmatic responses in a rat model of bronchial asthma. They found that REM sleep is related to nocturnal asthma, probably because of changes in sympathetic modulation. Nunes et al. [9] examined the effect of sleep deprivation on allergen-induced airway inflammation in an ovalbumin (OVA)-sensitized mice model and reported that there were severe lung inflammation in the sleep-deprived OVA-sensitized mice group compared to the control group. In detail, their results indicated that sleep deprivation is related to elevated IL-17 levels, pulmonary neutrophilia, and resistance to corticosteroid therapy. However, in this study, there were no significant differences in serum IL-1β and the number of inflammatory cells in BALF except for corticosterone between the two groups. These results are attributable to several limitations. It is mainly thought to be because of the small sample size and the period of sleep deprivation. Jung et al. [11] assessed if auditory functions such as hearing levels are influenced by the sleep deprivation in an animal model and established that sleep deprivation may induce increases of inflammatory cytokines and the destruction of cochlea hair cells, causing hearing loss. In their study, a significant elevation in IL-1β levels was found in the sleep deprivation nine days group (n = 12) compared to the control group (n = 15), in future studies, it is necessary to use more samples or to apply varied sleep deprivation periods. Our result showed that there was a significant increase in the corticosterone levels in the SD group compared to the HS group. These outcomes are consistent with those of many types of research that estimate serum corticosterone concentrations in rats with sleep deprivation. Choi et al. [12] investigated the association between sleep deprivation and the male reproductive system, including blood steroid hormone levels, and found that corticosterone levels significantly elevated in the sleep deprivation in the four days and seven days groups compared to the control group.
One of the main disadvantages of this study was that the number of animals used in this experiment was too small (only three rats per group). In the modified platform model, we used the rats live for 72 hours by moving between platforms. However, if we have too many rats in a platform model, the cage is too crowded, and rats cannot move freely, which is another stressor. In our previous research, we found that if too many rats were placed in the platform model, there could be various problems such as drowning. Since this study has the characteristics of a pilot study, in the future, we will endeavor to confirm the results of this study and deduce more meaningful conclusions by repeating experiments several times.
Previous studies show that stress can increase airway inflammation [13]. The mechanism may be the endogenous circulating corticosterone significantly modulate immune cell differentiation and function. For example, a stress-induced hormone promotes pro-allergic Th2-type responses and the level of corticosterone in plasma inhibits the IL-12/interferon-γ regulatory axis [14-16].
In conclusion, it is crucial to reveal the association between sleep and allergy in modern society, where sleep duration is gradually decreasing, and allergic diseases are increasing. The results of this pilot study indicate that sleep loss may be associated with deterioration of allergic inflammatory response. Further study using more subjects is required.